光是調控植物生長發育的重要環境信號。植物種子在土壤中發芽後,通過暗形態建成促進幼苗快速伸長,出土見光;然後通過光受體介導的光信號轉導促進光形態建成,來抑製下胚軸伸長、促進子葉和葉片的擴展以及葉綠素的合成,使幼苗得以進行光合作用👩💼,從而實現自養生長🧀。bHLH類轉錄因子PIFs(包括PIF1🌨👓、PIF3、PIF4和PIF5)是紅光/遠紅光受體光敏色素(phytochromes)的關鍵下遊因子,通過促進下胚軸的伸長來抑製光形態建成過程。紅光下激活的光敏色素B (phyB)通過與PIFs互作👩❤️👩,誘導其磷酸化和蛋白降解,從而促進光形態建成👩🏿🦳☄️。H2A.Z作為一種組蛋白變體🪼,在植物溫度感知🕜、發育和開花時間調控等過程中發揮重要作用;SWR1復合體則負責催化H2A.Z在染色質中的占位。目前為止✌🏿,已知PIF4和SWR1復合體在調控溫度依賴的植物生長過程中具有十分緊密的關系,但是二者發揮作用的生化分子機製尚不明確。
4月28日⛏,Journal of Genetics and Genomics在線發表我校生命科學學院楊洪全教授和茅誌磊副研究員團隊題為“PIFs interact with SWC6 to regulate H2A.Z deposition and photomorphogenesis in Arabidopsis”的研究論文🤹🏽♀️。該研究發現擬南芥光敏色素互作因子PIFs通過與SWR1復合體亞基SWC6直接互作🤍,抑製SWC6與光形態建成關鍵正調控因子HY5的結合,減少H2A.Z在生長素響應基因上的占位,從而抑製光形態建成🍔。

該研究首先發現在酵母細胞和體外系統中PIF1、PIF3🚣🏿♀️、PIF4和PIF5分別與SWC6存在互作🌿;在煙草細胞共表達PIF1🐅、PIF3👩👩👧、PIF4和PIF5分別與SWC6的兩兩組合,在細胞核中可定位到相同的核斑點🥙;免疫共沉澱實驗顯示PIF3和PIF4與SWC6在擬南芥體內存在互作。通過把swc6和arp6分別與pifq四突變體雜交,得到swc6 pifq和arp6 pifq五突變體,比較紅光下下胚軸長度,發現SWC6和ARP6在紅光下主要通過依賴於PIFs來控製下胚軸的伸長🕺。基因表達結果顯示PIFs與SWC6和ARP6拮抗地調控生長素響應基因IAA6☝️、IAA19、IAA20以及IAA29的表達🤽🏿♂️;ChIP-qPCR結果顯示pifq突變體中在IAA6和IAA19基因上的H2A.Z占位水平比WT明顯上升。通過蛋白體外和體內實驗🥪,發現PIFs可能通過與HY5競爭結合SWC6來抑製SWR1復合體在這些基因上的催化活性,從而調控H2A.Z的占位和光形態建成。
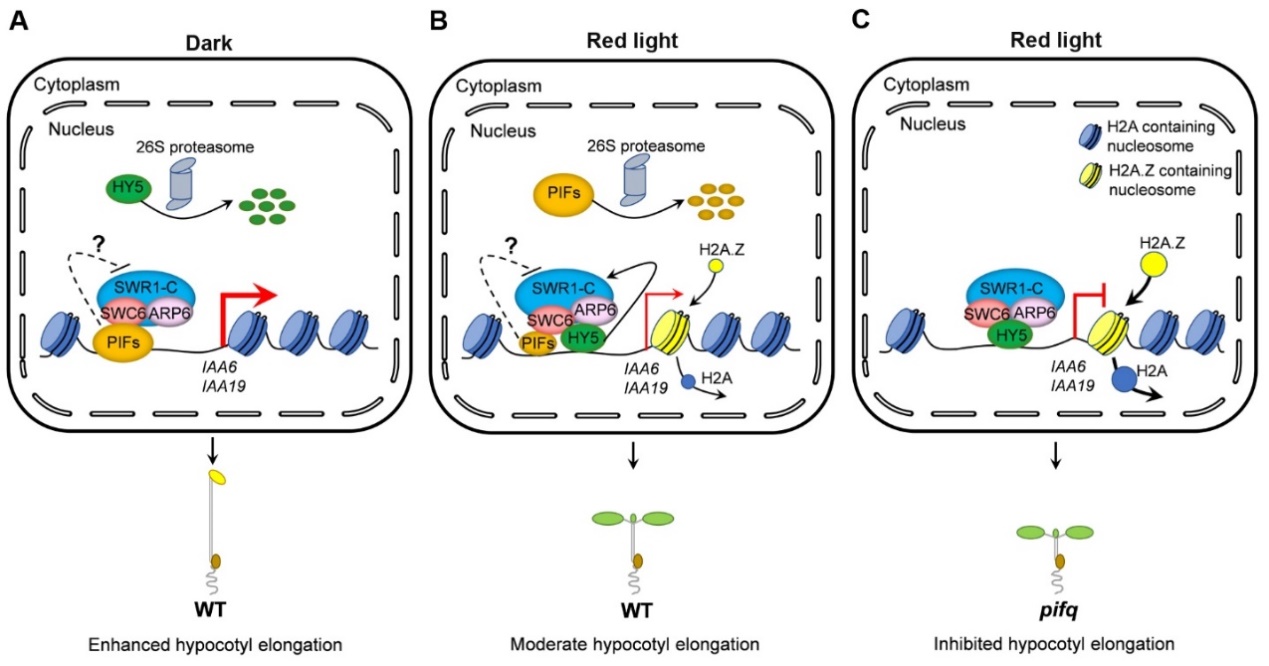
PIFs參與H2A.Z在生長素響應基因IAA6, IAA19占位的調控機製
A:在黑暗生長的野生型幼苗中,PIFs大量積累,而HY5通過泛素化26S蛋白酶體途徑發生降解🤽🏻。因此,PIFs與SWC6互作,充分抑製SWR1復合體的活性和H2A.Z的占位,進而充分激活IAA6和IAA19的表達,促進下胚軸伸長。
B:在紅光下生長的野生型幼苗中🏊🏼♂️,HY5蛋白大量積累👩🏿🦰,而PIFs因先後發生磷酸化泛素化和蛋白降解導致其蛋白水平較低。因此,HY5與SWC6和ARP6的互作只能在一定程度上被低水平的PIFs所抑製,使SWR1復合體以合適的活性來促進H2A.Z在IAA6和IAA19基因上占位😐,從而維持這些基因表達處於合適的水平以及適度的下胚軸伸長。
C🚳:在紅光下生長的pifq突變體幼苗中🦿🐃,由於沒有PIFs蛋白🤦,HY5與SWC6和ARP6的互作不受PIFs的抑製,使SWR1復合體能充分促進H2A.Z在IAA6和 IAA19基因上的占位👨❤️👨,從而抑製這些基因的表達和下胚軸的伸長。
綜上🧕🏿,該研究發現PIFs通過與SWC6的互作,抑製H2A.Z在生長素響應基因上的占位來促進這些基因的表達,進而調控光形態建成的分子機製🧑🏼🦰。678体育平台博士生陳慧茹為該論文第一作者🧍🏻♀️,茅誌磊副研究員為通訊作者。該研究得到國家自然科學基金、國家重點研發計劃等項目的資助。
論文鏈接:https://www.sciencedirect.com/science/article/pii/S1673852723000954?via%3Dihub
